Mechanism of Collective Cell Migration
The movement of cells is central to a range of normal and disease processes, including embryonic and tissue development, tissue repair and wound healing, angiogenesis, cancer invasion and metastasis, immune function and inflammation. In multicellular organisms, certain cells, such as leukocytes and fibroblasts, tend to move as single individuals, while others, such as epithelial and endothelial cells, generally migrate as groups, maintaining cell-cell contacts as they move.
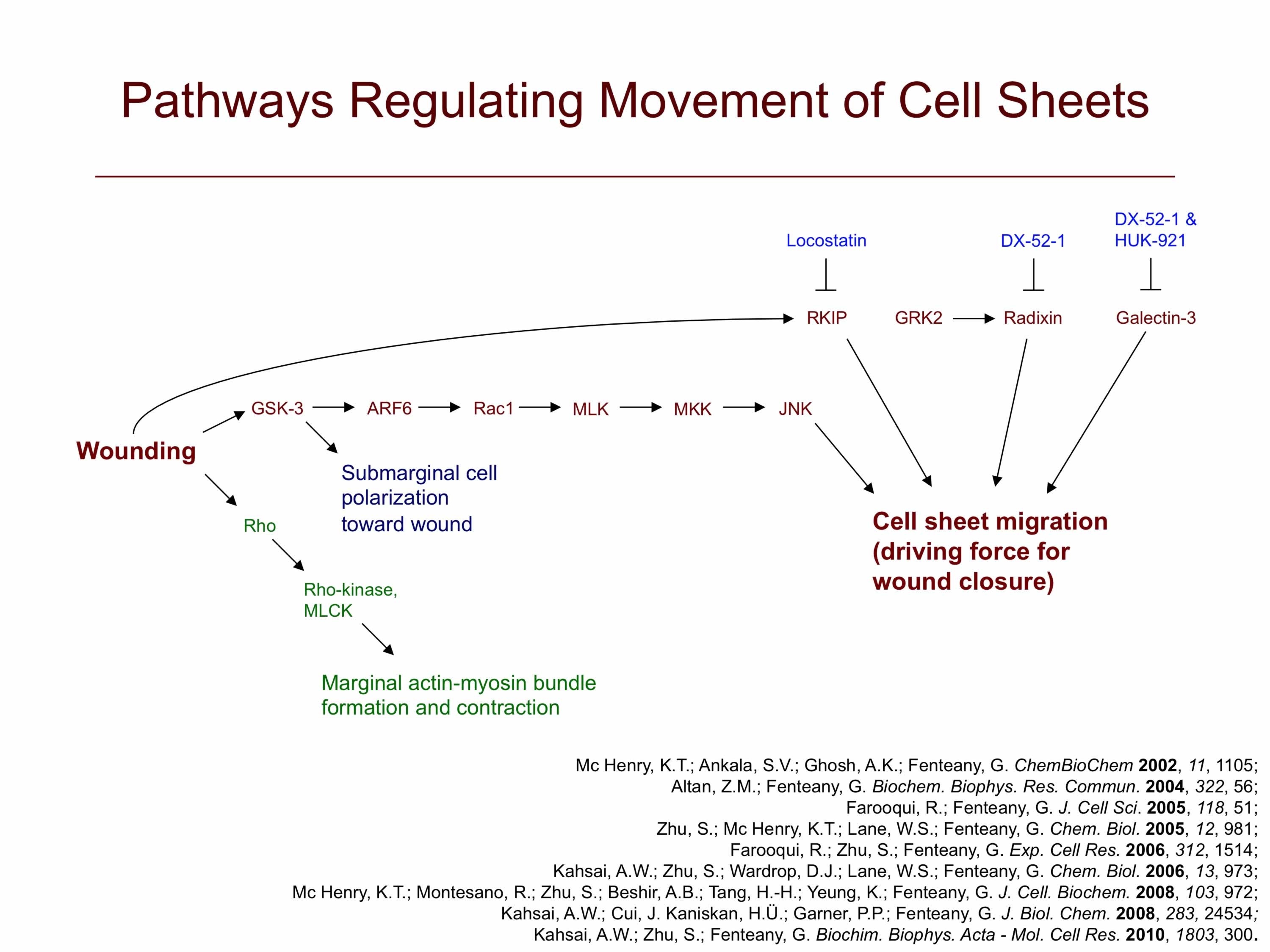
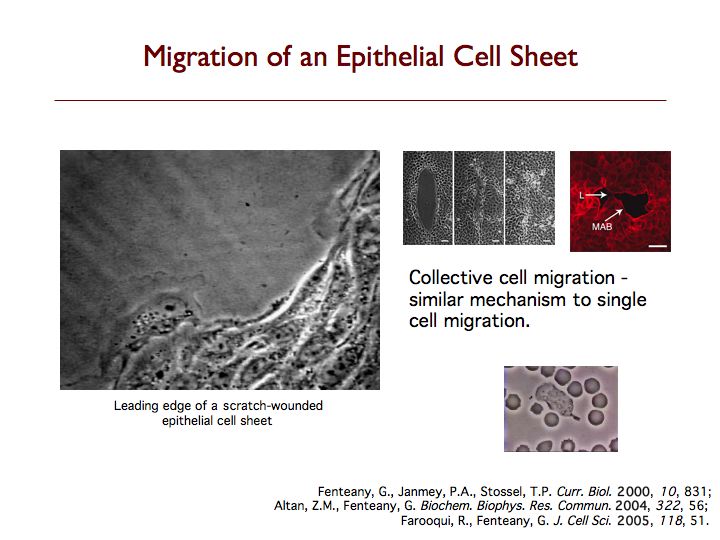
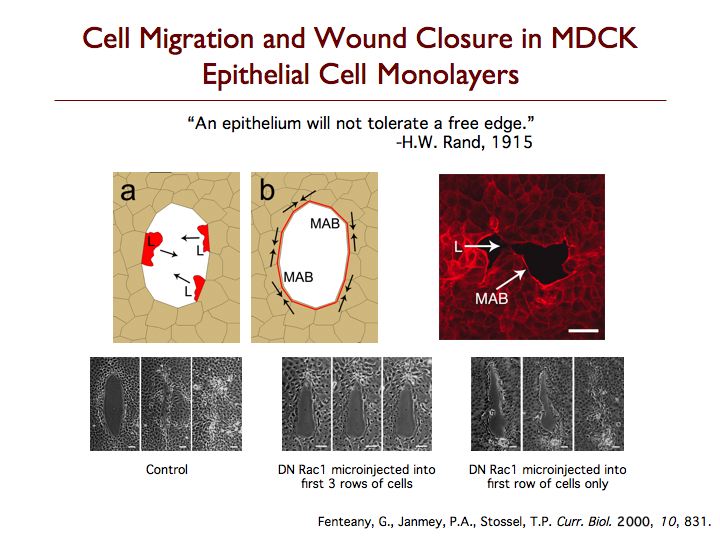
The mechanism by which epithelial, endothelial and other strongly cell-cell adhesive cells migrate collectively as continuous sheets is not clear. We have found that wounds in Madin-Darby canine kidney (MDCK) cell sheets close by protrusive cell crawling involving the small GTPase Rac, phosphoinositides and active movement of multiple rows of cells. Wounds induced in MDCK epithelial cell monolayers close by Rac- and phosphoinositide-dependent cell crawling, with formation of lamellipodia at the wound margin, and not by contraction of a perimarginal actomyosin purse-string. Although Rho-dependent actin bundles usually form at the margin, neither Rho GTPase activity nor formation of these structures is required for wound closure to occur at a normal rate. In fact, the marginal actin bundles appear to slightly antagonize protrusion and migration while making the advancement of the wound edge slightly more even and regular, possibly by helping to distribute force from more actively protruding cells to neighboring cells at the margin. Cdc42 GTPase activity is also not required for closure. Inhibition of Rho or Cdc42 results, however, in statistically significant decreases in the regularity of wound closure, as determined by the ratio of wound margin perimeter over the remaining denuded area at different times. The Rac-dependent force generation for closure is distributed over several rows of cells from the wound margin, as inhibition of motility in the first row of cells alone does not inhibit closure and can be compensated for by generation of motile force in cells behind the margin. Furthermore, high levels of Rac-dependent actin assembly in the first few rows of cells from the wound margin are observed. These findings suggest a new distributed mode of signaling and movement that, nevertheless, resembles individual cell motility. Although Rho and Cdc42 activities are not required for closure, they have a role in determining the regularity of closure. MDCK cell sheet migration also depends upon the function of c-Jun N-terminal kinase, glycogen synthase kinase-3 and Raf kinase inhibitor protein.
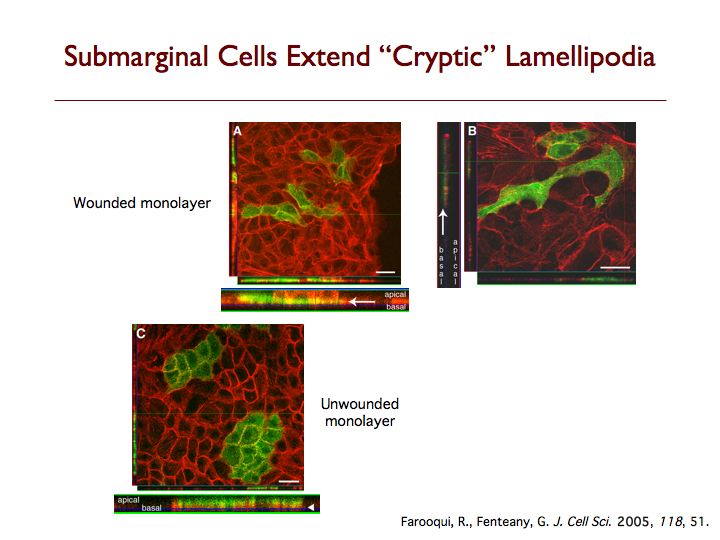
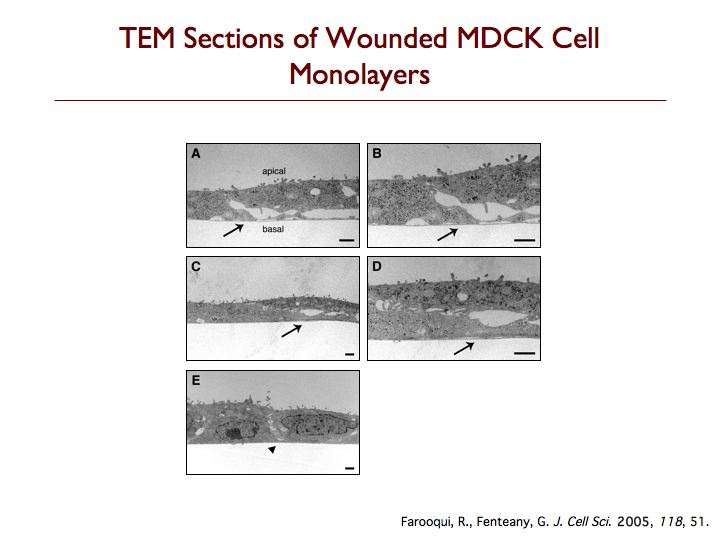
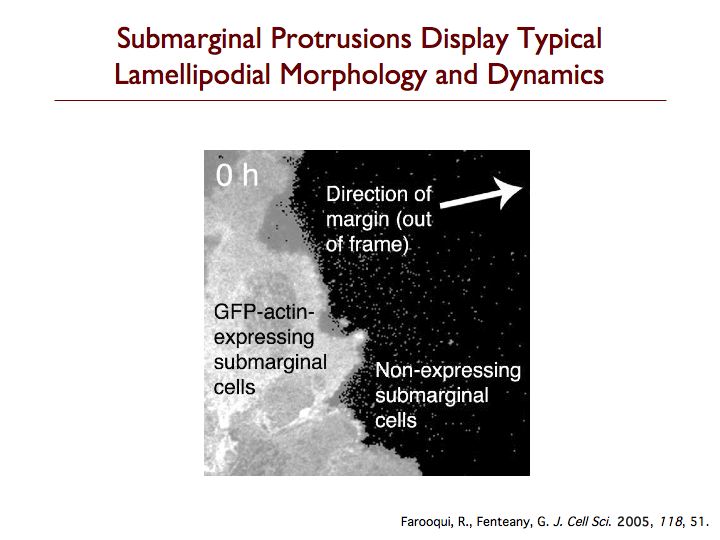
We have discovered that cells behind the margin of wounded MDCK cell monolayers, even hundreds of microns from the edge, extend “cryptic” lamellipodia against the substratum beneath cells in front of them toward the wound, as determined by confocal, two-photon and transmission electron microscopy. These so-called submarginal cells nevertheless strictly maintain their more apical cell-cell contacts as they migrate as part of a coherent cell sheet, hiding their basal protrusions from conventional microscopy. The submarginal protrusions display the hallmarks of traditional lamellipodia based on morphology and dynamics. Cells behind the margin therefore actively crawl, instead of just moving passively as cells at the margin pull on them. The rate of migration is inversely proportional to distance from the margin, and cells move coordinately yet still partly autonomously toward the wound area. In addition, some cell proliferation occurs at a delay after wounding but does not contribute to closure, instead apparently serving to replace damaged cells so that intact spread cells can revert to their normal cuboidal morphology and the original cell density of the unbroken sheet can be restored. We are now exploring the basic mechanism of cell sheet migration in more detail and working on the discovery of new small-molecule probes of cell motility and the elucidation of their mode of action.
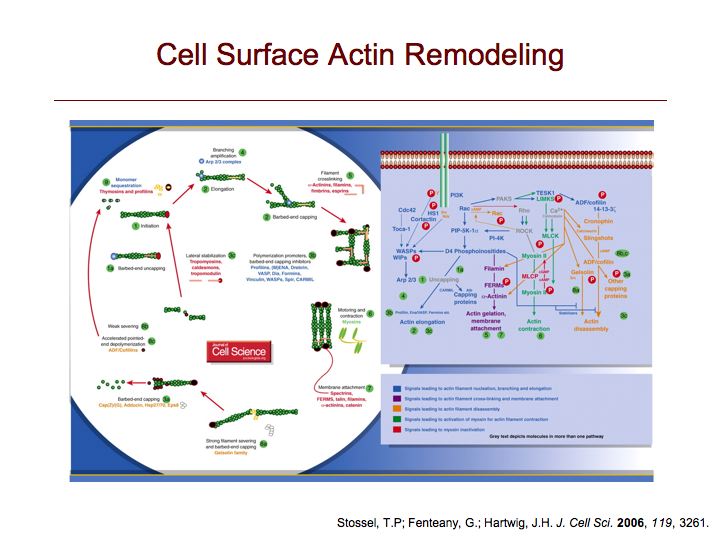
More on signaling pathways to cell motility.